Results/Discussion
I set out to validate the lichen cover-to-biomass conversion factors I used in my study by doing destructive lichen sampling as described in the Methods section. Figure 12 compares the McMullin conversion factors to the conversion factors I developed. A series of two-tailed T-tests for each pair of conversion factors indicated that my conversion factors were not significantly (α = 0.05) different from McMullin's (p > 0.40).
I set out to validate the lichen cover-to-biomass conversion factors I used in my study by doing destructive lichen sampling as described in the Methods section. Figure 12 compares the McMullin conversion factors to the conversion factors I developed. A series of two-tailed T-tests for each pair of conversion factors indicated that my conversion factors were not significantly (α = 0.05) different from McMullin's (p > 0.40).
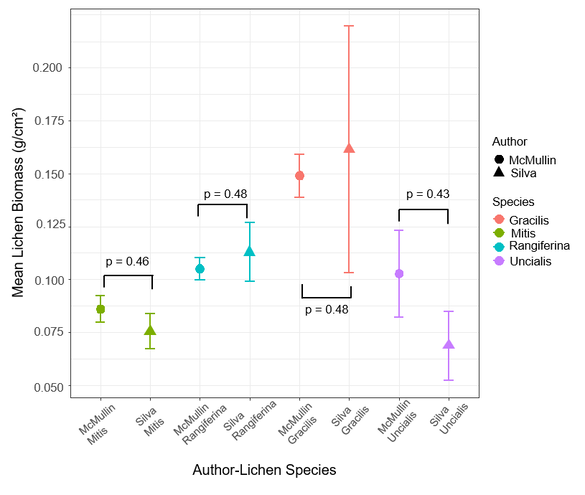
Figure 12. Comparison of cover-to-biomass conversion factors developed by McMullin et al. (2011) and Joe Silva for four lichen species eaten by woodland caribou. Error bars represent ± 1 SE.
The boxplots in Figure 13 demonstrate post-fire lichen recovery in my study area at the transect level. I grouped the transects by decadal burn classes, with the exception of stands > 50 years old. I decided to combine the older burn classes by creating a class for burns 50-70 years old and a class for burns 70-120 years old. This helped address the small sample sizes of the older age classes. Virtually no lichen was present in stands that burned within the last 20 years (Figure 13). No clear trend in lichen recovery was observed in dense conifer stands, indicating this ecosite is unlikely to support lichen foraging habitat for caribou. On the contrary, starting 20 years post-fire, sparse conifer demonstrated a marked increase in lichen biomass decade-over-decade. Lichen biomass peaked at a median of 3,090 kg/ha in sparse conifer stands 50-70 years post-fire.
The boxplots in Figure 13 demonstrate post-fire lichen recovery in my study area at the transect level. I grouped the transects by decadal burn classes, with the exception of stands > 50 years old. I decided to combine the older burn classes by creating a class for burns 50-70 years old and a class for burns 70-120 years old. This helped address the small sample sizes of the older age classes. Virtually no lichen was present in stands that burned within the last 20 years (Figure 13). No clear trend in lichen recovery was observed in dense conifer stands, indicating this ecosite is unlikely to support lichen foraging habitat for caribou. On the contrary, starting 20 years post-fire, sparse conifer demonstrated a marked increase in lichen biomass decade-over-decade. Lichen biomass peaked at a median of 3,090 kg/ha in sparse conifer stands 50-70 years post-fire.
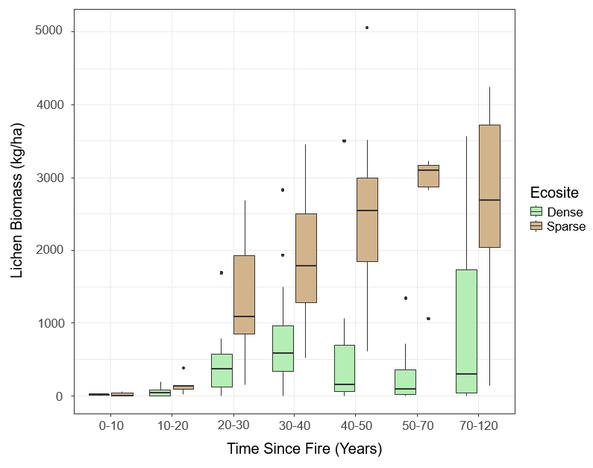
Figure 13. Lichen biomass (kg/ha) for seven burn age classes in the study area.
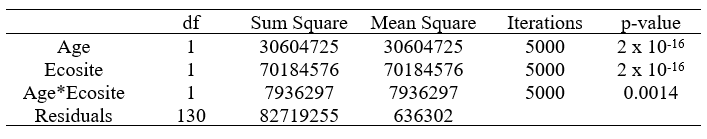
I ran a permutational ANOVA to determine if there is a significant relationship between time since fire and lichen biomass. The p-values in Table 5 suggest there are significant (α = 0.05) relationships between age and lichen biomass, and ecosite and lichen biomass. The output also indicates there is a significant interaction between age and ecosite, likely due to the divergent post-fire recovery trajectories of sparse conifer and dense conifer, as noted in Figure 13.
Table 5. Permutational ANOVA table for Lichen Biomass ~ Age*Ecosite.
I ran a permutational ANOVA to determine if there is a significant relationship between time since fire and lichen biomass. The p-values in Table 5 suggest there are significant (α = 0.05) relationships between age and lichen biomass, and ecosite and lichen biomass. The output also indicates there is a significant interaction between age and ecosite, likely due to the divergent post-fire recovery trajectories of sparse conifer and dense conifer, as noted in Figure 13.
Table 5. Permutational ANOVA table for Lichen Biomass ~ Age*Ecosite.
I used piecewise regressions of lichen biomass and age to estimate a breakpoint for each ecosite. I interpreted the breakpoint as the post-fire age at which lichen biomass had recovered. The piecewise regression for sparse conifer revealed a significant relationship between age and lichen biomass (p = 8.76 x 10-9), with age explaining 48% of the variation in lichen biomass (Figure 14). The breakpoint was estimated at 49 years, with a standard error of ± 7.5 years, which is consistent with Environment Canada's 40-year recovery threshold. The two lines in the piecewise regression demonstrate that lichen biomass increased quite rapidly (62 kg/ha/yr) for the first 49 years after a fire, but subsequently remained virtually constant.
I used piecewise regressions of lichen biomass and age to estimate a breakpoint for each ecosite. I interpreted the breakpoint as the post-fire age at which lichen biomass had recovered. The piecewise regression for sparse conifer revealed a significant relationship between age and lichen biomass (p = 8.76 x 10-9), with age explaining 48% of the variation in lichen biomass (Figure 14). The breakpoint was estimated at 49 years, with a standard error of ± 7.5 years, which is consistent with Environment Canada's 40-year recovery threshold. The two lines in the piecewise regression demonstrate that lichen biomass increased quite rapidly (62 kg/ha/yr) for the first 49 years after a fire, but subsequently remained virtually constant.
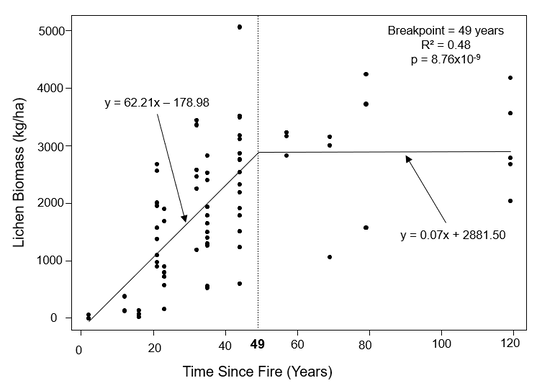
Figure 14. Piecewise regression for sparse conifer transects (n = 74). The breakpoint indicates that lichen has recovered 49 years after fire.
The piecewise regression for dense conifer sites indicated there is no significant relationship between lichen biomass and age (p = 0.12), with age only explaining 14% of the variation in lichen biomass (Figure 15). This validates my observation that lichen biomass never really recovers in dense conifer stands after fire, demonstrating that these sites are unlikely to support terrestrial forage lichens for woodland caribou. The estimated breakpoint for dense conifer stands is 31 years, with a standard error of ± 10.6 years. Similar to sparse conifer stands, lichens begin to recolonize dense conifer about 20 years after fire, but quickly occupy all suitable microsites, reaching an early breakpoint and remaining virtually constant thereafter.
The piecewise regression for dense conifer sites indicated there is no significant relationship between lichen biomass and age (p = 0.12), with age only explaining 14% of the variation in lichen biomass (Figure 15). This validates my observation that lichen biomass never really recovers in dense conifer stands after fire, demonstrating that these sites are unlikely to support terrestrial forage lichens for woodland caribou. The estimated breakpoint for dense conifer stands is 31 years, with a standard error of ± 10.6 years. Similar to sparse conifer stands, lichens begin to recolonize dense conifer about 20 years after fire, but quickly occupy all suitable microsites, reaching an early breakpoint and remaining virtually constant thereafter.
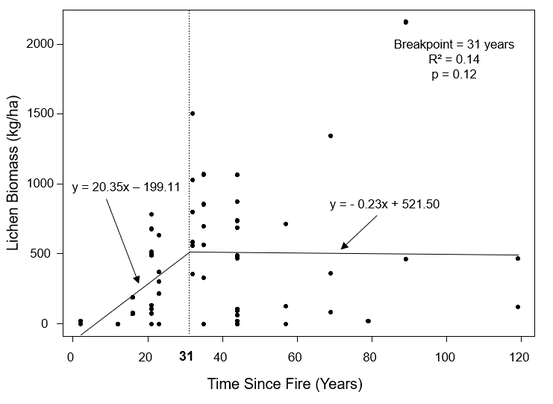
Figure 15. Piecewise regression for dense conifer transects (n = 60). The breakpoint indicates that lichen biomass has recovered 31 years after fire.
I used generalized linear mixed models to determine which set of variables best predicts lichen abundance, and ranked the models using AIC. Both model sets (i.e.: lichen biomass vs. lichen cover as the response variable) predicted the same top model (Canopy/Litter), which included ecosite, age, canopy closure and litter as predictor variables. Upon closer inspection of the model outputs, the Canopy/Litter model for lichen cover had much smaller standard errors for the coefficients, and the residual plots indicated better fit between the predicted and observed values compared to the corresponding model for lichen biomass. Thus, AIC scores and associated statistics are only reported for the lichen cover models (Table 6).
Table 6. Ranking candidate models using Akaike's Information Criterion (AIC). K indicates the number of fixed effects in the model, ∆AIC the change in AIC, and wi the Akaike weight for each model.
I used generalized linear mixed models to determine which set of variables best predicts lichen abundance, and ranked the models using AIC. Both model sets (i.e.: lichen biomass vs. lichen cover as the response variable) predicted the same top model (Canopy/Litter), which included ecosite, age, canopy closure and litter as predictor variables. Upon closer inspection of the model outputs, the Canopy/Litter model for lichen cover had much smaller standard errors for the coefficients, and the residual plots indicated better fit between the predicted and observed values compared to the corresponding model for lichen biomass. Thus, AIC scores and associated statistics are only reported for the lichen cover models (Table 6).
Table 6. Ranking candidate models using Akaike's Information Criterion (AIC). K indicates the number of fixed effects in the model, ∆AIC the change in AIC, and wi the Akaike weight for each model.
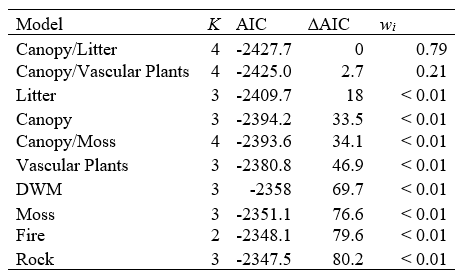
Models with a ∆AIC > 2 are considered significantly less informative than the model with the lower score (Harrison et al. 2018). In this case, the Canopy/Vascular Plants model has a ∆AIC > 2, indicating the Canopy/Litter model is the best of the set. This is reflected in the high Akaike weight (wi = 0.79) which indicates the likelihood the Canopy/Litter model is the best of the set. Based on the coefficient estimates for the Canopy/Litter model (Table 7), sparse conifer ecosites have a positive influence on lichen cover. Age has a positive effect on lichen cover, whereas canopy closure and litter both have negative effects on lichen cover.
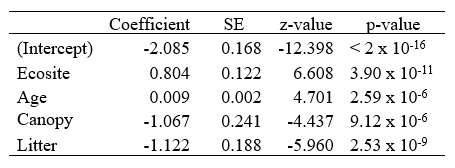
Table 7. Coefficient table for the Canopy/Litter model with lichen cover as the response variable.